
Intens trening aktiverer proteinsyntese, men bare hvis riktig ernæring er der for å støtte den. Hvis du leser artikkelen John Meadows og jeg skrev kalt Maximize Protein Synthesis, eller hvis du har lest noe som T Nation har sagt om emnet i årevis, er du sannsynligvis kjent med konseptet "det anabole vinduet" og viktigheten. av peri-trening ernæring.
Det som skjer på mobilnivå i timene etter trening er forutsigende for langsiktige gevinster. Dra nytte av dette "anabole vinduet", og du vil vokse som aldri før. Gå glipp av det hele tiden, og vel, lykke til.
Å få de rette makronæringsstoffene til rett tid er nøkkelen, men makroer er bare en del av det store bildet. Et viktig, men ofte oversett aspekt av muskelproteinsyntese, er cellevolum. Cellevolum er ikke bare kosmetisk; det er den viktigste driveren for aminosyretransport, og jobber også bak kulissene for å slå på proteinsyntese og undertrykke proteinnedbrytning.
En full / volumisert muskel er en anabole muskel. Mens vi har kjent i over 20 år at hevelse av celler hemmer proteinnedbrytning og stimulerer proteinsyntese i visse celler (1-3), men inntil nylig var mekanismen som knytter cellevolum til proteinsyntese et mysterium.
Det vi vet nå er at proteinsyntese styres av enzymet mTOR, som aktiveres av mekanisk stress, vekstfaktorer og leucin.
Selv om alle disse tre er viktige for treningsstimulansen, er mTOR-signalering også avhengig av cellevolum.(4) Dette er spesielt viktig i skjelettmuskulaturen, der cellevolumisering aktiverer glykogen- og proteinsyntese og hemmer proteinnedbrytning.(5, 6)
Det vitenskapelige gjennombruddet som førte til sammenhengen mellom cellevolum og proteinsyntese skjedde i 2005, da en gruppe forskere oppdaget at det kreves mer enn bare leucin for å aktivere mTOR - glutamin er også nødvendig.(7)
Dette kom som en overraskelse. Selv om glutamin betraktes som en "betinget essensiell" aminosyre som begrenser proteinnedbrytning under alvorlig traume eller stress, hadde det aldri vært knyttet til mTOR-aktivering.
Glutamin var nødvendig for leucinopptak og cellevolumisering, som begge er nødvendige for å aktivere proteinsyntese. Forfatterne fortsatte med å vise det cellulære glutaminet utarmning resulterer ikke bare i en reduksjon av cellevolumet, men reduserer også leucins evne til å aktivere proteinsyntese.(7)
Denne oppdagelsen var enorm fordi den ga en direkte sammenheng mellom glutamin, cellevolumisering og proteinsyntese. For første gang ble det vist at glutamin er nødvendig for aktivering av proteinsyntese med leucin.
Mens denne studien antydet at glutamin er en veldig viktig del av puslespillet som knytter cellevolum og proteinsyntese, ble den eksakte mekanismen ikke utarbeidet før 2009, da Nicklin et al. oppdaget at glutamineksport er koblet til leucinimport og mTOR-aktivering.(8)
For å få leucin inn i cellen, er det en første periode med "glutaminbelastning.”Dette trekker også inn vann, og øker cellevolumet. Etter "glutaminbelastningsfasen" eksporteres glutamin ut av cellen i bytte for import av leucin.
Nicklin et al. oppdaget også at cellulære glutaminnivåer er hastighetsbegrensende for aktivering av proteinsyntese av leucin. Når celler ble behandlet samtidig med glutamin og en EAA-blanding som inneholder leucin, ble aktivering av proteinsyntese forsinket i 60 minutter. Når de samme cellene ble "forhåndslastet" med glutamin, ble proteinsyntese slått på innen 1-2 minutter etter at cellene ble gitt leucin.
Dette resultatet var viktig fordi det forklarte forsinkelsestiden for aktivering av proteinsyntese av leucin i denne eksperimentelle modellen.
Disse resultatene kaster endelig lys over mobilmaskineriet som regulerer aminosyretransporten, og hvordan den er koblet til kontrollen av proteinsyntese.
Dette arbeidet må tolkes med en viss forsiktighet. En viktig advarsel for disse studiene var at de ble utført in vitro (i.e. i cellekulturer) der regulering eller proteinsyntese er mye enklere. Muskelceller er i stand til å lage glutamin etter behov fra andre aminosyrer, og "glutaminutarmning" i denne cellekulturmodellen er ikke representativ for mer fysiologiske situasjoner in vivo.
Høye proteinsyntese kan ikke opprettholdes på ubestemt tid i muskelvev med eller uten glutamintilskudd. Glutamin kan imidlertid brukes til strategisk å støtte proteinsyntese ved å optimalisere cellevolumisering i løpet av perioden etter trening.

Celler er veldig opptatt, og det er mange membranbundne ionekanaler og transportørproteiner som regulerer trafikk inn og ut av cellen. Det er spesielt to klasser av aminosyretransportører som er viktige her: "System L" og "System A" aminosyretransportører er tettest knyttet til mTOR-signalering og proteinsyntese.(8-10)
Aktiviteten til System A og System L-transportører er koblet, noe som gjør at leucin og de andre BCAAene kan absorberes i cellen.(11) System L-transportører er ansvarlige for tilstrømningen av leucin og de andre BCAAene i bytte mot utstrømningen av glutamin.
System A-transportører arbeider imidlertid gjennom en annen mekanisme der glutamin er koblet til natriumopptak.(12, 13) Koblingen mellom natriumopptak og System L / System A aminosyretransportører kalles Tertiær aktiv transport (forkortet TAT). Det er TAT som til slutt driver leucin inne i cellen, noe som fører til mTOR-aktivering og proteinsyntese.(11)
Du kan se hvordan TAT fungerer i figuren nedenfor:
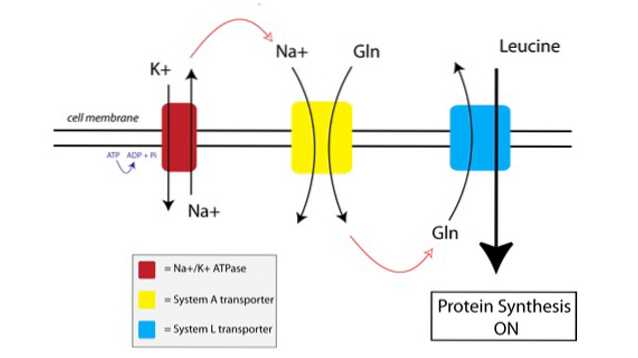
For det første bruker en membranbundet pumpe kalt natrium-kalium ATPase-pumpe (Na + / K + ATPase, rød i figuren ovenfor) energi fra ATP for å flytte natrium utenfor cellen, mot konsentrasjonsgradienten.
Den økte natriumkonsentrasjonen utenfor cellen er koblet til importen av glutamin av System A-transportøren (gul på figuren). Glutamin og natriumtilstrømning til cellen trekker også inn ekstra vann, noe som får cellen til å hovne opp. Dette setter cellen i anabole tilstand, og primer proteinsyntesemaskineriet for aktivering.
Når glutamin bygger seg til tilstrekkelig høye nivåer inne i cellen, aktiveres System L-transportørene (blå på figuren), som skyter glutamin utenfor cellen i bytte for leucinopptak. Innføring av leucin i cellen er utløseren for proteinsyntese.
Selv om dette har vært en god biokjemieleksjon så langt, er oppdagelsen av TAT ikke bare viktig for cellebiologer. Nå som vi vet hvordan cellevolum er koblet til aminosyretransport og proteinsyntese, kan vi designe flere ernæringsstrategier for å maksimere prosessen når den teller - i den kritiske perioden etter trening.
Proteinsyntese er helt avhengig av cellehydrering - hvis du til og med er litt dehydrert, er evnen til å komme seg etter intens trening fullstendig kompromittert. Å få rikelig med vann er ikke noe bra her, men vann alene er ikke nok.
Elektrolytter som natrium, kalium, klorid og fosfat fungerer også som "osmolytter" fordi de trekker vann inn i cellen. Etter en intens treningsøkt, trenger vi vann, aminosyrer og elektrolytter for å maksimere cellevolumiseringsprosessen som driver proteinsyntese.
Natrium, magnesium, kalsium, kalium, fosfat og klorid (for å nevne noen) er alle viktige her. På et minimalt nivå (med mindre du gjør det etter råd fra legen din), ikke vær vekk fra natrium før eller etter trening. Hvis du er utarmet av natrium, vil pumpen din nesten ikke eksistere, og natrium er nødvendig for opptak av glutamin.
For å eliminere gjetningen er Surge® Workout Fuel og Plazma ™ designet med ideelle elektrolyttforhold for å støtte cellevolum og proteinsyntese.
Glutaminopptak i cellen forårsaker cellevolumisering, og primer muskelceller for proteinsyntese. Som nevnt er en full / volumisert muskel en anabole muskel. Sammen med å drive aminosyretransport, øker cellevolumisering også glykogensyntese og hemmer proteinnedbrytning.(4-6)
Proteinsyntese undertrykkes av utarmning av glutamin, som har enorme implikasjoner for idrettsutøvere med hard trening. Etter en intens treningsøkt er en inflammatorisk respons montert, som gjør at immunceller kan komme seg inn i myket muskelvev for å starte reparasjons- / gjenoppbyggingsprosessen.(14)
Glutamin tas så raskt opp av immunceller at det regnes som “drivstoffet i immunsystemet.”(15) Ikke overraskende er det vist at intens trening forårsaker plasmaglutaminutarmning.(16-18)
Av denne grunn øker kravene til glutamin i perioden etter trening, hvor den lokale immunresponsen kan konkurrere om tilgjengeligheten av glutamin til primære muskelceller for opptak av aminosyrer og proteinsyntese.
Forhåndslading celler med glutamin kan også potensielt redusere "lag-time" assosiert med leucinaktivering av proteinsyntese. Hvis du ikke allerede gjør dette, ta 10-15 g glutamin eller glutaminpeptider umiddelbart etter trening. Fordi BCAA er et annet yndlingssubstrat for muskelglutaminsyntese, og det også har vist seg å øke muskelglutaminproduksjonen (19-21), er BCAA og leucin også nyttige i løpet av perioden før trening for å bidra til å maksimere endogen glutaminproduksjon.
Redaktørens merknad: Hele Plamza / MAG-10-protokollen, med ernæring før trening, trening og etter trening, oppfyller forfatterens anbefalinger for å laste musklene med BCAA og leucin.
Nylig ble det oppdaget at forbruket av EAA øker uttrykket for både System A og System L aminosyretransportører.(9) Det er viktig at dette skjer på "post-transkripsjonsnivå", som betyr på nivået med proteinsyntese, hvor eksisterende mRNA blir oversatt til proteiner.
Kontraster dette med “denovo”Proteinuttrykk - der det kan ta 16 eller flere timer å syntetisere, behandle og transportere nye mRNAer - aktivering av proteinsyntese etter transkripsjon kan forekomme i løpet av minutter til timer, slik at celler raskt kan øke nivået av bestemte proteiner når det er nødvendig.
Plutselig har vi mer insentiv til å ha en solid ernæringsplan for peri-trening - EAA-inntak i pre- og peri-treningsperioden lønner seg stort etter trening ved å øke uttrykket for aminosyretransportører, og primere cellen for maksimalt aminosyreopptak og aktivering av proteinsyntese.
Rasktabsorberende proteinisolater eller hydrolysater som Mag-10® Protein Pulsing Protocol ™ eller Plazma ™ i løpet av pre- og peri-treningstidene er ideelle her.
Insulin er det mest anabole hormonet i kroppen. Sammen med direkte aktivering av proteinsyntese, øker insulin også translokasjon av System A-aminosyretransportører til cellemembranen.(22)
Dette betyr at insulin får flere System A-transportører til å vises på cellemembranen, klare til å kjøre mer glutamin inn i cellen. Mer glutamin fører til mer cellevolum, som driver mer leucin inn i cellen, og til slutt fører til mer proteinsyntese.
Mens EAA øker uttrykk av AA-transportører, er det insulinsignalet som gjør at de kan vises på celleoverflaten, klare til å skyve nye aminosyrer inn i cellen.
Dette er enda en grunn til at karbohydrater før og peri trening er en god ide med mindre du er i ekstrem fett tapsmodus: insulin øker kapasiteten for cellulær aminosyretransport.
Karbohydrater øker insulinnivået, men visse aminosyrer kan også brukes til å styrke insulinfrigivelsen. Glutamin er en kraftig aktivator av "inkretin" hormoner, som gjør insulinproduserende celler i bukspyttkjertelen mer følsomme for glukose.(23) Glycin styrker også insulinfrigivelse gjennom en annen mekanisme.
Selv om karbohydrater etter trening alene vil øke insulinnivået, vil kombinasjon av disse insulinpotensierende aminosyrene med karbohydrater overbelaste bukspyttkjertelen for enda mer frigjøring av insulin. Selv om det er bra å holde insulinnivået på undersiden mesteparten av tiden, maksimerer insulinnivået i peri-treningsperioden maksimalt aminosyretransport, cellevolum og proteinsyntese, samtidig som proteinunderskudd undertrykkes.
Den typen intens anaerob trening som trengs for å bygge massevis av kvalitetsmuskler fører til betydelig laktatproduksjon, redusert pH i muskler. Dette fører til tidlig muskeltretthet og svakhet, men også visse aminosyretransportører, inkludert System A, hemmes av lav pH.(1. 3)
Når pH i muskler er lav, reduseres aminosyreopptaket, noe som undertrykker mTOR-aktivering av proteinsyntese.(24) Det er også vist at lav pH-inhibering av System A-aminosyretransportører øker proteinnedbrytningen.(25)
Dette er hvor beta-alanin kommer inn. Økte muskelkarnosinnivåer fungerer som en naturlig syrebuffer, og utvider den anaerobe terskelen ved å begrense reduksjonen i muskel-pH fra trening.
Beta-alanin har en annen viktig funksjon: å bidra til å opprettholde proteinsyntese og få den raskere online etter intens trening ved å forhindre demping av aminosyretransport.
For å øke karnosinnivået i muskler, ta to tabletter Beta-7 ™ tre ganger om dagen.
Under intense treningsøkter reduseres proteinsyntese og proteinnedbrytning aktiveres. Dette er uunngåelig for enhver hardtreningsløfter. I hvor stor grad vi kan minimere de katabolske effektene av trening, og jo raskere vi kan gå tilbake til "anabole modus" i løpet av perioden etter trening, avgjør imidlertid til slutt hvor effektivt vi vil komme oss - og vokse.
Makronæringstiming er viktig, men det er et middel til et mål. Cellevolum er den viktigste driveren for aminosyretransport og proteinsyntese. Ved å forstå hvordan aminosyretransport skjer, og hvordan det reguleres av cellevolum, kan vi få mer leucin raskere til søppel i muskelceller, og dermed føre til anabole brannen og til slutt føre til bedre gevinster.
Ovennevnte strategier er effektive, praktiske og er basert på den nyeste vitenskapelige forskningen. Bruk dem som en mal for å ta treningsfremdriften til neste nivå.
Jeg ser frem til spørsmålene dine på LiveSpill!



Ingen har kommentert denne artikkelen ennå.